“A non-mendelian pattern of inheritance governed by the DNA present in the cytoplasm is known as extrachromosomal inheritance or cytoplasmic inheritance.”
Our genetic material- DNA has arranged on chromosomes. It stores, transfer and express genetic information through replication, transcription and translation. Nuclear DNA- present in the nucleus of a cell, governs all phenotypes and is transmitted from parents to offspring in a specific pattern.
Autosomal dominant, autosomal recessive, X-linked dominant and X-linked recessive are several well-known patterns of inheritance. Genes present on autosomes follow autosomal inheritance whilst genes present on the X chromosome follow X-linked inheritance.
Broadly, when genotypes transfer from parents to offspring through reproduction are categorized in Mendelian inheritance. Nonetheless, several genes present in the cytoplasm don’t follow it. The process is known as cytoplasmic inheritance, non-Mendelian inheritance or extrachromosomal inheritance.
Organelle like chloroplast or mitochondria possesses cytoplasmic genes and these strings of DNA are often referred to as organelle DNAs.
In the present article, I am focusing on extrachromosomal inheritance, how it occurs and several examples. I will also explain maternal inheritance and the criteria of how it occurs. This present article will be a great learning opportunity for students regarding the present topic.
Let’s start with the basics,
Key Topics:
What is extrachromosomal inheritance?
The extrachromosomal inheritance also known as cytoplasmic inheritance or non-mendelian inheritance was first reported by Boris Ephrussi in yeast in 1949. As we said, organelles like chloroplast or mitochondria have their own genome. However, scientists still don’t understand how these organelles have created their own genomes.
One theory stated that it was a symbiotic relationship. It is believed that mitochondria were once free-living bacteria. Over a period of time, it has created a symbiotic relationship with eukaryotic cells and established itself in the cytoplasm. And later on, evolved as an organelle.
Similarly, the chloroplast in green plants comes from the free-living algae, established a symbiotic relationship with eukaryotic plant cells and settled into the cytoplasm of green plants. If you wish to learn about the present topic, we have given a link above in the article.
Both types of sub-genomes have well-developed DNA machinery. They can do their own replication, transcription and translation. In addition, the chloroplast has an antibiotic resistance gene which indicates that it has evolved from bacteria, previously.
The extrachromosomal genome comprises a few genes of several thousand base pairs, mostly are encoding rRNA, tRNA and other proteins for their DNA metabolism. Note that they have their own inheritance pattern.
Definition:
“The DNA present in the cytoplasm and not on chromosomes which follows the non-Mendelian pattern of inheritance is known as extrachromosomal inheritance.”
Now in the next section, I am going to explain about criteria, what are the criteria to categorize it as extrachromosomal DNA.
Criteria for extrachromosomal inheritance:
The extrachromosomal DNA follows a non-mendelian pattern of inheritance
Unlike the common nuclear DNA, genes present on organelle don’t follow the Mendelian style of Inheritance. It is a circular genome, lacking centromere and that’s why can’t segregate like the genomic DNA.
Their own machinery for protein synthesis
As these sub-genome has their own machinery for replication, transcription and translation, it synthesizes their own DNA and makes their own protein.
Maternal inheritance:
The extrachromosomal DNA is inherited from the maternal side.
The segregation is observed in somatic cells rather than germ cells, unlike nuclear inheritance.
Examples:
Carl Correns in 1908, first reported non-mendelian inheritance in Mirabilis Jalapa plastid DNA. Another extrachromosomal inheritance was reported by M M. Rhoades in 1933. He postulated that the inheritance of male sterility in maize is governed by maternal inheritance and it becomes one of the greatest discoveries in science.
Another important point that makes extrachromosomal DNA even unique is maternal inheritance. Meaning, it transmits from mother to offspring. This shows that female individuals of the entire population are capable of transferring the cytoplasmic DNA.
No scientific evidence exists explaining why and how it occurs, however, One theory suggests that the female reproductive cell (ovum) is bigger than the sperm cell, contains more cytoplasm and more organelles than a male reproductive cell. Researchers expecting that this influences non-mendelian inheritance or maternal inheritance.
One of the classical examples of maternal inheritance is,
Cytoplasmic male sterility in maize.
In maize, nuclear genes do not play any significant role rather, the sterility is inherited through the egg cytoplasm in offspring.
When a male sterile plant is crossed with a normal fertile plant, all the F1 plants remain sterile. When all F1 sterile plants are backcross with a normal fertile plant, until all chromosomes from the male sterile line are exchanged to male fertile, the sterility persists in the progeny.
The image below makes it clear,
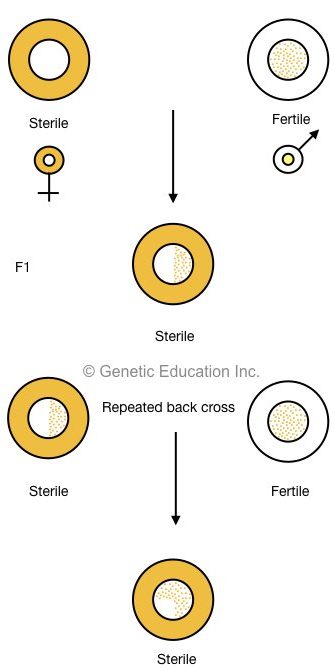
Generally, male-sterile lines are denoted as tcs, T (Texas), C (Cytoplasmic), S (Sterility). It was believed that T (Texas) cytoplasm is associated with susceptibility against several types of disease like leaf blight disease and yellow blight disease in maize.
Results indicating that chromosomal/nuclear DNA does not significantly affect male sterility (particularly in maize). Furthermore, most of the cytoplasm, carrying organelles are inherited from the maternal side. present explanation confirms that the sterility transmits from the cytoplasm.
Present findings become an important milestone in plant research and crop improvement. Maize corns can be uniformly developed in hybrid sterile maize plants.
Maternal inheritance is a miracle in nature, how some genotypes govern by the maternal side influences several important phenotypes. However, nuclear/chromosomal inheritance also occurs from the maternal side. Materna inheritance in complex, studies shows that several genotypes from the maternal side also influence phenotypes.
Read some of the interesting articles:
The maternal-effect in snail:
The character of coiling in snail is governed by maternal inheritance. Snail- Limnaea peregra, has two types of shell coiling phenotypes: one is dextral that coil for the right side and another is sinistral that coils for the left side.
Here, the mother’s genotype (not a phenotype) is exclusively responsible for the development of coiling style. Assume that D+ genotype codes for dextral (right side) coiling and D codes for sinistral coiling. The reciprocal cross of D+ and D is shown in the figure:
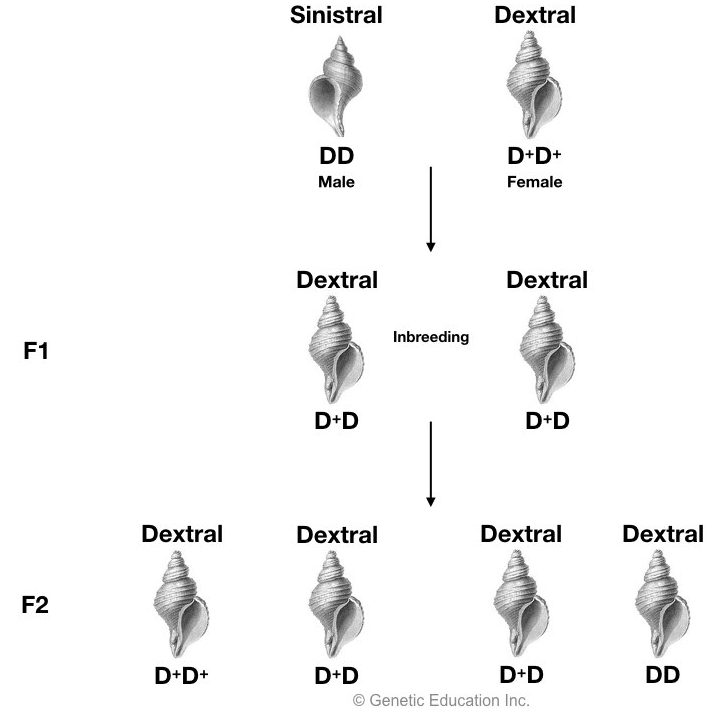
When crossing between D+D+ female and DD male, all the F1 and F2 progeny become dextral as the mother is D+D+ dextral Here, the DD recessive phenotype is not expressed and typical Mendelian 3:1 ratio is not obtained (all four are dextral).
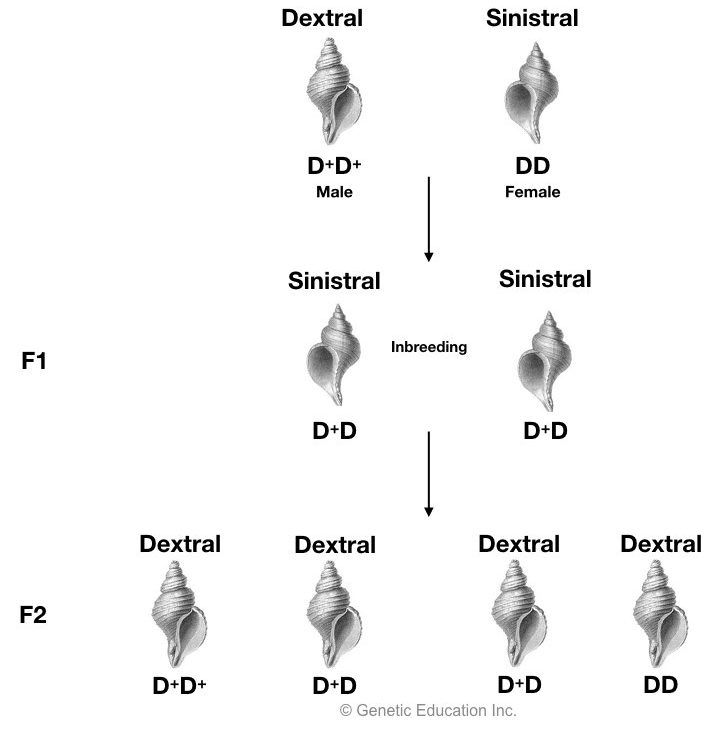
In another condition when DD sinistral female is crossed with D+D+ dextral male, F1 offspring become sinistral with genotype D+D, here mentioning genotype is important because the inheritance is governed by genotype not by phenotype.
When this F1 progeny is inbred (D+D * D+D) all the F2 progeny becomes dextral and coil for the right side. These results indicated that the phenotype of parents does not have any influence on the phenotype of progeny because although all of the F1 progeny are sinistral, all F2 offspring becomes dextral.
Detailed investigation shows that spindle formed during the second metaphase division decides the direction of coiling. The spindles of the dextral snail are tipped to the right and vice versa for sinistral. Interestingly, spindle arrangement in metaphase is controlled by maternal genes.
So the actual phenotype of “type of coiling” in snails is governed by maternal genes and it does not depend on the phenotype of any parents.
Interestingly in another scenario, the amount of cytoplasm transferred play a role in deciding the phenotype.
 Inheritance of kappa particles in paramecium:
Inheritance of kappa particles in paramecium:
“Paramecin” is a substance found in some of the killer strains of paramecium which kill the sensitive strains. Paramecin production is governed by the kappa particles present in the cytoplasm of the paramecium.
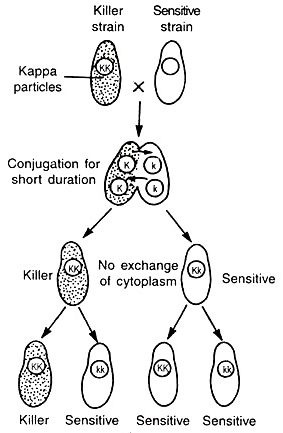
Here KK gene is responsible for the production of kappa particles which is dominant over the kk gene. In the case of inheritance of kappa particles, cytoplasmic exchange during conjugation plays a crucial role.
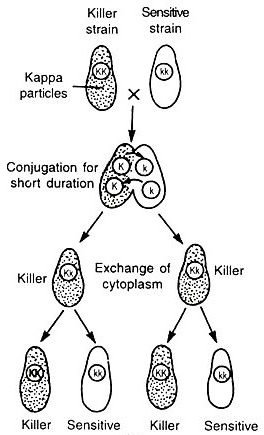
When KK killer strains are crossed with kk strains by conjugation, all the progeny obtained are heterozygous with genotype Kk but the phenotype of paramecium depends on the presence or absence of kappa particles and it will be influenced by the time of conjugation.
If both are conjugates for a shorter period of time, in the F1 generation killer strains remain killer and non-killer remain non-killer in the heterozygous condition. Here nuclear genes are only inherited and not cytoplasmic.
In another condition, if killer and non-killer strains are conjugated for a longer period of time, due to the exchange of kappa particles, sensitive strain receives kappa particles through cytoplasmic exchange and sensitive strains become killer in the F1 generation.
Extrachromosomal inheritance PDF, slide share and ppt:
Here I am enlisting some of the external resources that you can use for study. The PDFs, slide shares and ppts listed here are given from the trusted sources.
Two best slide share and ppt options for extrachromosomal inheritance:
Conclusion
The cytoplasm is an important component of the cell, not only for transferring organelles but also for the inheritance of characters. Cytoplasmic inheritance and maternal inheritance are responsible for some of the disease condition in the human. Any defect in the inheritance of extrachromosomal genes results in serious physical, mental and biochemical problems.
The next section of this topic comprises information about the organelle DNAs. You can read it here: Extrachromosomal inheritance class 2: organelle DNA.
Article was written by- Tasneem Gandhi
The article reviewed by- Tushar Chauhan
This article is written by our dearest friend Tasneem Gandhi on the special request of team genetic education. We are heartily thankful to Tasneem.